The Saccadic System - Pulse-Step Generator#
Learning Outcomes#
Learning goals
The saccadic system rapidly and precisely redirects the line of sight toward a newly selected target. In this module, we focus on the brainstem circuitry that converts high-level movement commands into the actual eye movement, through a coordinated interaction of burst neurons, pause neurons, integrators, and motoneuron pools.
After completing this module, you will be able to:
Describe and Draw the Main Sequence and identify its key features (duration–amplitude relationship and saturating peak velocity).
Explain why these features imply that the saccadic system is nonlinear
Contrast those features with the predictions of a linear system (constant duration and linear velocity scaling).
Draw the linear main sequence
Explain why a purely step-like motoneuron command cannot generate normal saccades and why a pulse–step pattern is required.
Describe the oculomotor plant as a second-order overdamped system, including its typical time constants and impulse response.
Explain how the plant’s mechanical properties constrain eye movement dynamics.
Identify the key brainstem nuclei responsible for horizontal saccades: EBNs, IBNs, LLBNs, OPNs, NPH, and oculomotor nuclei (nIII, nVI).
Explain the functional roles of:
EBNs and IBNs as the pulse generator
NPH as the neural integrator
OPNs as the saccade gate (fixation vs. movement)
LLBNs as the computation of instantaneous motor error
Describe the functional distinction between caudal Superior Colliculus (movement) and rostral Superior Colliculus (fixation), and how their interactions with OPNs trigger or prevent saccades.
Explain why internal negative feedback is essential for accurate saccades
Describe the evidence for internal motor error monitoring (e.g., interrupted saccades).
Describe the concept of motor error, how it evolves during the saccade, and how it drives burst neuron activity.
Describe the saturating nonlinear input–output relationship of burst neurons
Explain the traditional view that this nonlinearity was thought to produce the nonlinear features of the Main Sequence (e.g., peak velocity saturation).
Interpret the basic components of a computational saccade model, including the SC input, LLBN motor error computation, burst nonlinearity, neural integrator, and plant.
Predict qualitatively how lesions to different brainstem structures affect saccade metrics (velocity, amplitude, duration, fixation stability).
Optional Material
Some sections of this chapter include dropdown boxes with background or additional information and downloadable MATLAB code. These are for the current module optional and meant for students who want to explore the material in more depth. You do not need to study these parts to understand the core concepts, of this module.
The chapter also includes dropdown boxes with hints and/or answers. These questions are not optional, and may pop up in the exam.
General Background#
Saccades (saccadic eye movements) are rapid, ballistic shifts of gaze that allow us to redirect the line of sight toward objects of interest in our visual environment. These eye movements are an essential aspect of visual processing, allowing us to efficiently scan our environment, read, and attend to moving objects. Although we are usually unaware of them, humans make approximately three saccades per second when scanning a scene or reading. These movements are essential because only the central fovea provides high-resolution vision; to inspect objects in detail, the eyes must be brought to the correct location as quickly and accurately as possible.
The saccadic system is one of the best-studied sensorimotor systems in primates. Its movements are fast, stereotyped, and highly reproducible, which makes it an ideal model system for understanding how the brain transforms sensory information into motor commands. Over decades of research, the oculomotor system has provided fundamental insights into neural coding, internal feedback, population dynamics, decision making, and motor control.
This module focuses on the brainstem oculomotor circuitry responsible for generating the actual saccade once a target has been selected. We begin by reviewing the characteristic properties of saccades — such as their duration, peak velocity, and the nonlinear “Main Sequence” — and relate these features to the mechanical properties of the oculomotor plant. We then examine how the brainstem and midbrain convert a desired eye movement into the appropriate pulse–step pattern that drives the extraocular muscles. This includes an introduction to the major neuronal populations involved in saccadic control: excitatory and inhibitory burst neurons (EBNs and IBNs), long-lead burst neurons (LLBNs), omni-pause neurons (OPNs), the neural integrator in the NPH, and the oculomotor nuclei.
Finally, we will develop and explore a biologically plausible computational model that simulates saccade generation. By linking model behavior to physiological constraints, we can understand why certain neuronal properties — such as pulse-like bursts, tonic firing, gating mechanisms, and internal feedback loops — are necessary for producing accurate and stable eye movements. This modeling framework also allows us to predict how specific neural lesions give rise to characteristic oculomotor deficits.
Saccades#
Saccade as a step response#
Saccades are rapid eye movements that shift the line of sight to a new visual target. From a Systems Theory perspective (Linear Systems), a saccade can be understood as the step response of the oculomotor system (Fig. 37). When a target suddenly appears at a new location, or when it “jumps” on a screen, the visual system experiences a step change in retinal error: the target’s position changes instantaneously, but the eyes have not yet moved.
The oculomotor system must therefore transform this step input — a sudden change in desired eye position — into a movement that brings the eyes to the new target. In an idealized linear system with infinite bandwidth, the output would instantly follow the input: the step response would be a perfect step. Biological systems, however, impose constraints:
Visual and motor processing introduce a reaction time before the eye starts to move.
The extraocular muscles and orbital tissues have inertia and viscosity, so the eyes cannot move instantaneously.
Perception and motor execution contain noise, meaning the end point of the movement may not land exactly at the desired location on every trial.
Despite these constraints, saccades come remarkably close to the ideal: they are extremely fast, highly accurate, and have short latencies. They represent the brain’s best possible approximation of a step under biological limitations. Later in this module, we will see that producing such an efficient “biological step” requires the brainstem to generate a characteristic pulse-step pattern of neural activity.
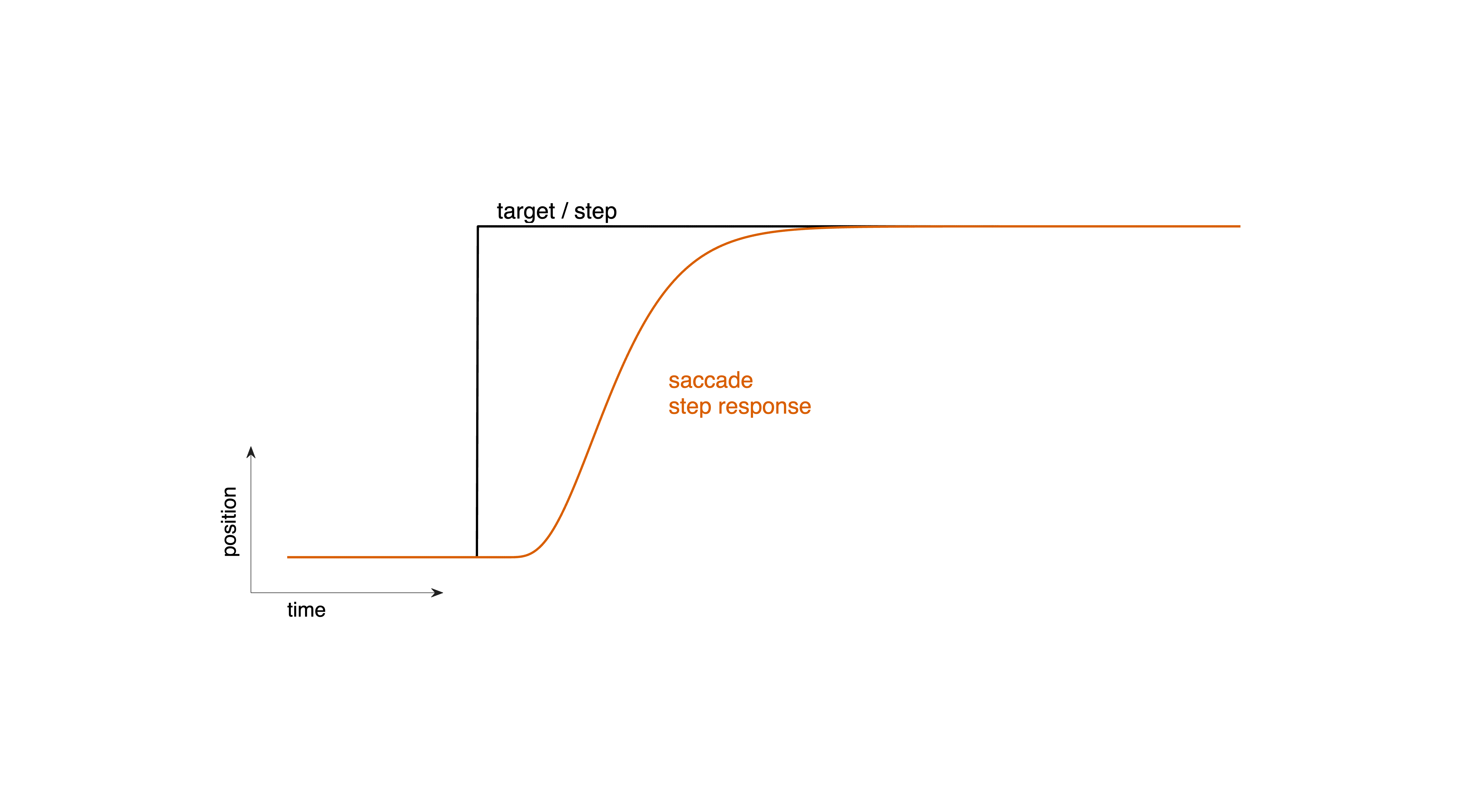
Fig. 37 | A saccade as a step response.#
The target jumps to a new location (step input), followed—after a brief reaction time—by a rapid shift of the eyes (the system’s step response).Download Matlab code to generate this figure
Main Sequence#
Saccades are among the fastest movements the body can produce, reaching peak velocities of up to 700 °/s in humans and 1300 °/s in rhesus monkeys. That saccades are fast is surprising. When a target appears unexpectedly, the resulting saccade shows a highly stereotyped temporal profile. Across individuals and species, saccades obey a characteristic set of relationships known as the main sequence (Fig. 38). The main sequence summarizes how three quantities co-vary:
Amplitude – the size of the eye movement
Duration – how long the movement lasts
Peak velocity – the maximum speed reached during the movement
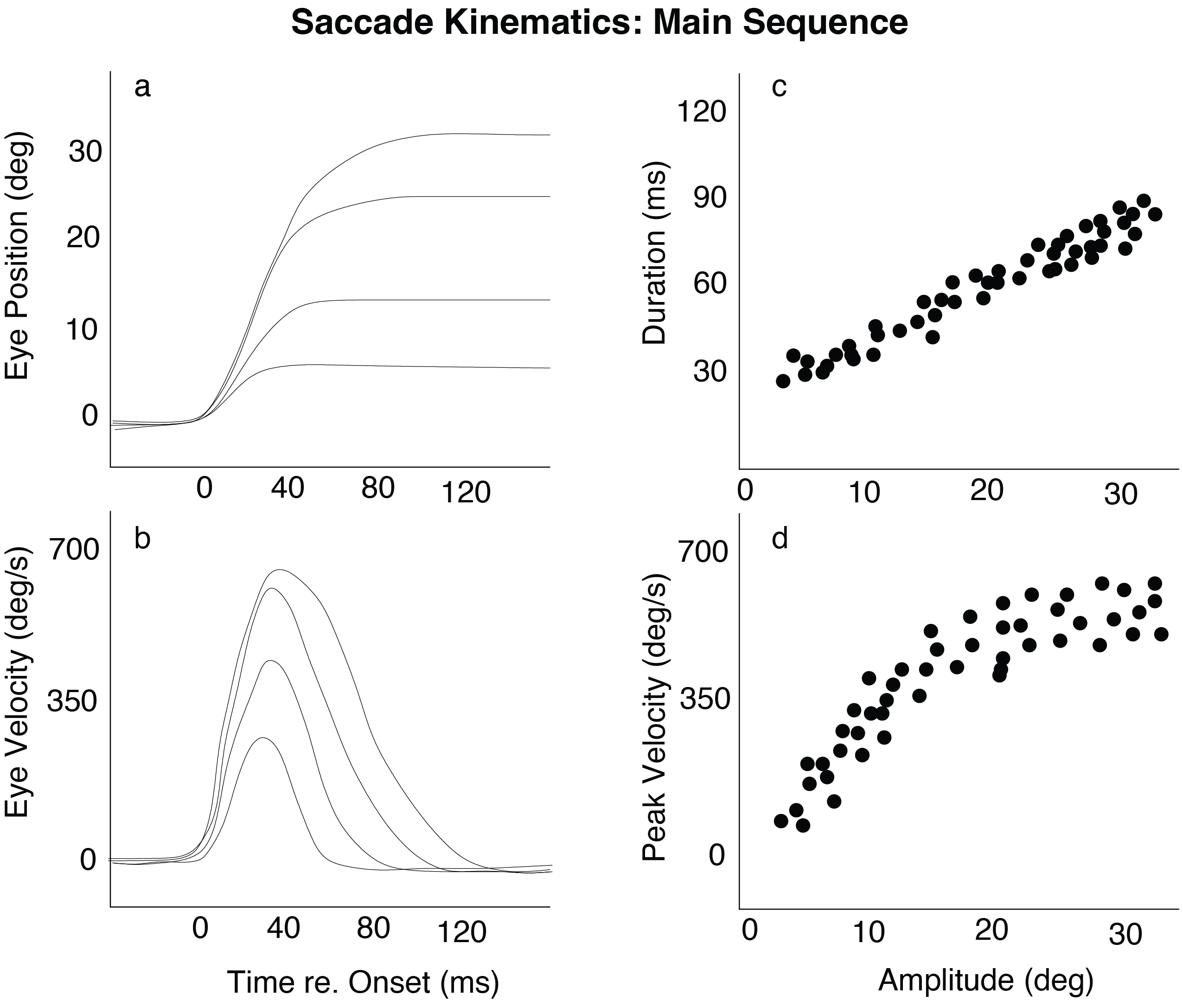
Fig. 38 | Saccadic kinematics for movements of varying amplitudes.#
a. Horizontal saccades as a function of time.
b. Velocity profiles for different amplitudes.
c. Duration increases approximately linearly with amplitude.
d. Peak velocity rises with amplitude but saturates.
The two classic Main Sequence plots are:
Duration vs. Amplitude (Fig. 38c) → roughly linear increase
Peak Velocity vs. Amplitude (Fig. 38d) → rapid rise followed by saturation
Because of this consistency, the main sequence has become a foundational tool in both research and clinical oculomotor assessment.
Neural Control#
One striking feature of saccades is how fast they are. The eyes and their supporting tissues — the extraocular muscles, the eyeball, connective tissue, and surrounding orbital mechanics, together called the oculomotor plant — form a mechanical system with inertia, viscosity, and elastic restoring forces. Intuitively, such a system should be relatively sluggish: if you were to push the eye with a simple, steady force, it would move only slowly and smoothly, not with the explosive speed seen in real saccades. This observation allows us to reason about the input the system must be receiving. Even before we know anything about brainstem neurophysiology, we can treat the eye-and-muscle system as a “black box” mechanical plant whose output is the observed saccade. From the kinematics alone — the main sequence — we can infer what kind of neural command signal must be driving the plant, and what kind of system can produce this control signal.
1. A simple step-like motoneuron command is insufficient#
If the oculomotor plant received a step command (a sudden change in tonic muscle activation), its movement would resemble the step response of a sluggish (overdamped second-order) system (Fig. 39): far too slow to produce real saccadic behavior.
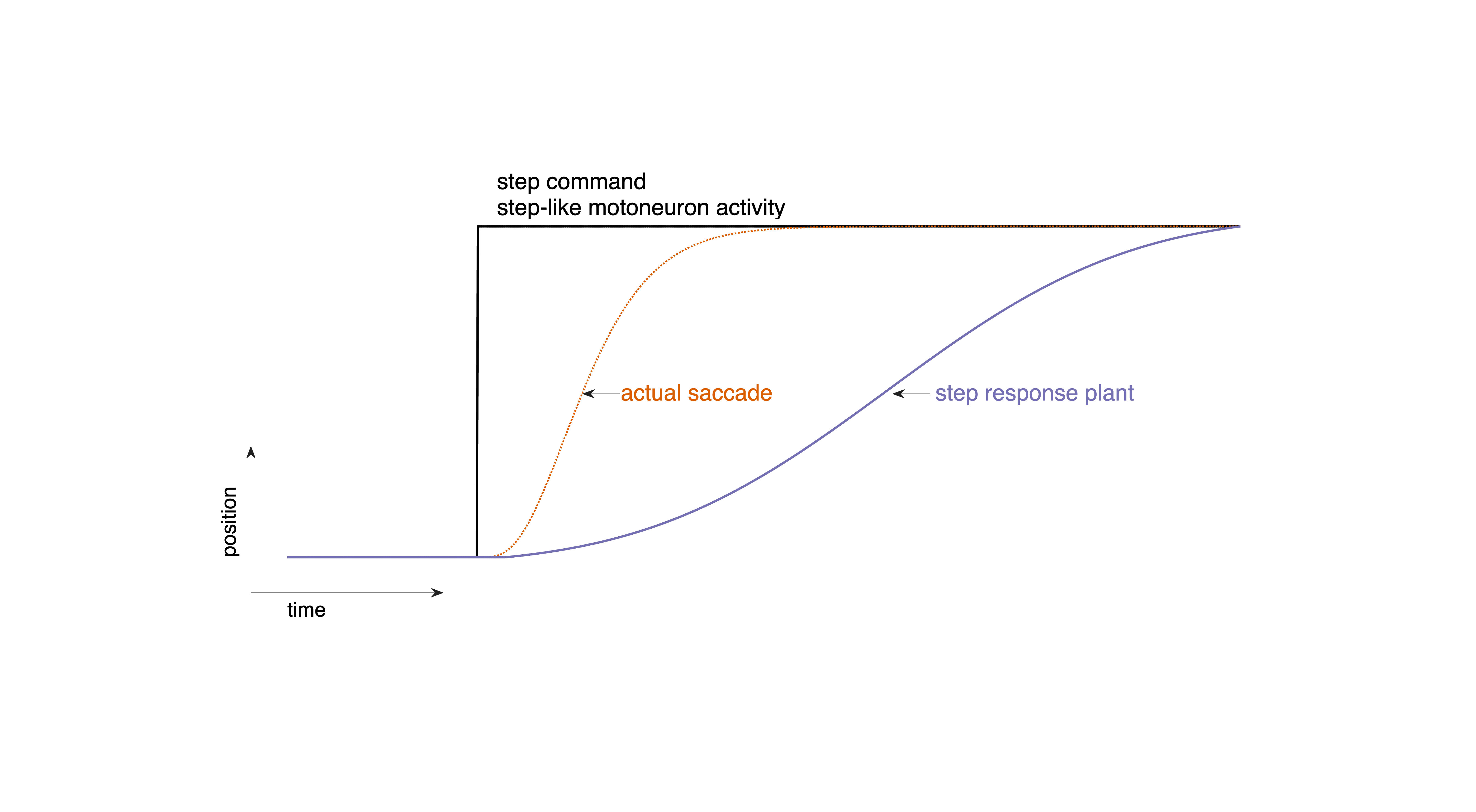
Fig. 39 | Step command#
A step command produces movements far too slow compared to real saccades. Download Matlab code to generate this figure
2. A pulse–step innervation is necessary#
If a steady step of neural activity cannot move the eyes fast enough, the plant must instead be driven by a brief, high-intensity burst of motoneuron activity: a pulse. Such a pulse provides a large, phasic (short-lived) force that rapidly accelerates the eyes to high velocities. This is exactly what we observe in real saccades: the eye shoots toward the target with an explosive onset, reaching peak velocities of hundreds of degrees per second within just a few milliseconds.
Once the eye reaches its new position, the high-frequency pulse must stop — otherwise the eye would overshoot. But the eye must stay at the new eccentric position, which requires a tonic level of activation to counteract the plant’s elastic restoring forces: a step in neural activity.
Thus, the saccadic control signal must have two components:
Pulse: a brief, high-amplitude burst that generates the rapid movement
Step: a lower, sustained firing rate that keeps the eye in its new position
Together, this pulse–step command (Fig. 40) produces the fast, accurate, and stable saccades observed in the Main Sequence.
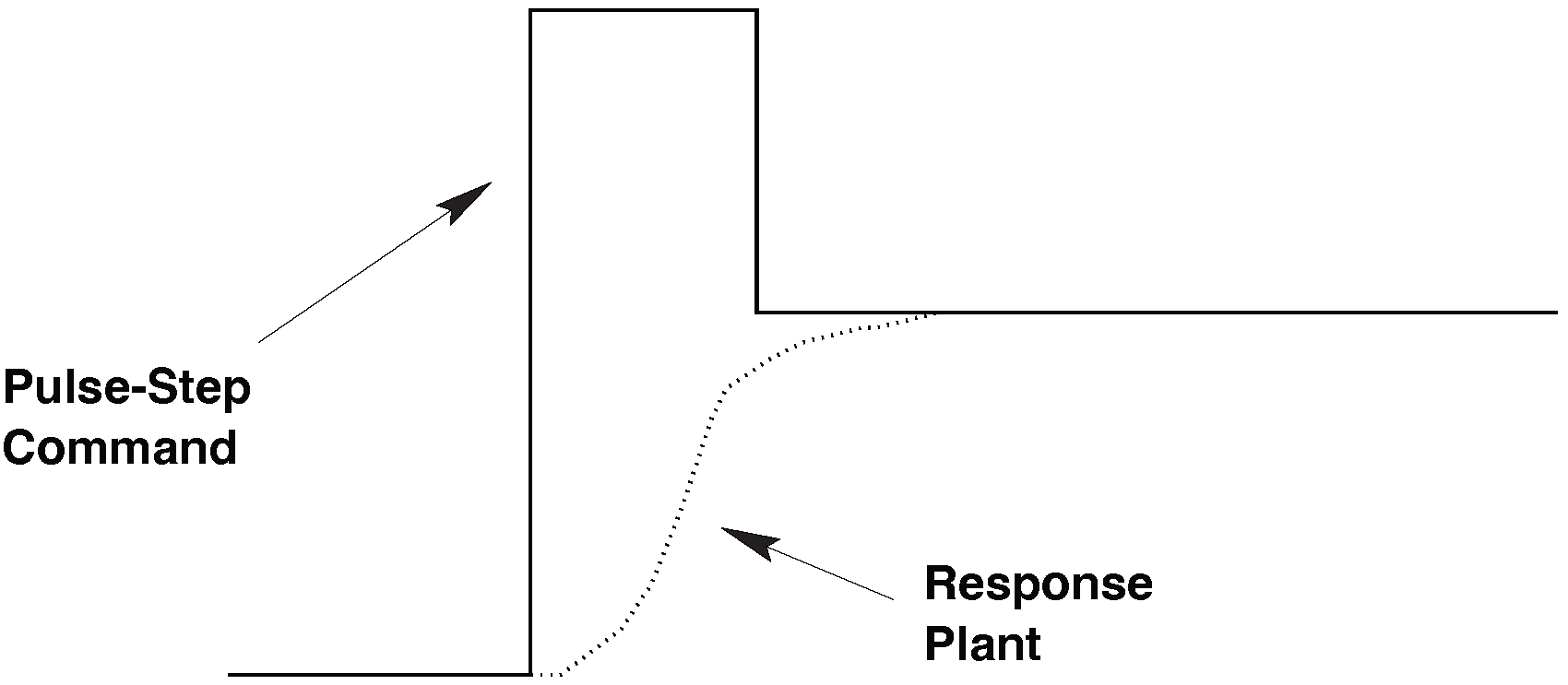
Fig. 40 | Pulse-step command.#
A pulse–step command produces realistic saccadic kinematics.
3. The saccadic system is nonlinear#
A linear system has the property that scaling the input by a factor \(a\) scales the output by the same factor (Fig. 41):
From this, a linear saccadic system would predict:
Constant duration for all amplitudes (a scaled response is simply taller, not longer).
Peak velocity increasing linearly with amplitude (no saturation).

Fig. 41 | Linear Saccadic System.#
Linearity requires proportional input–output scaling.
However, the actual Main Sequence (Fig. Fig. 38) violates both predictions:
Duration increases with amplitude.
Peak velocity saturates, instead of increasing proportionally.
These deviations clearly indicate that the saccadic system contains intrinsic nonlinear mechanisms. Later modules will explore where these nonlinearities originate (historically attributed to the burst generator, but now known to also arise upstream).
question – linear main sequence
Why would a linear system produce saccades with
(1) constant duration and
(2) peak velocity proportional to amplitude?
Sketch the Main Sequence you would expect for any linear system. See also the Hint/Answer in the next dropdown box.
Hint
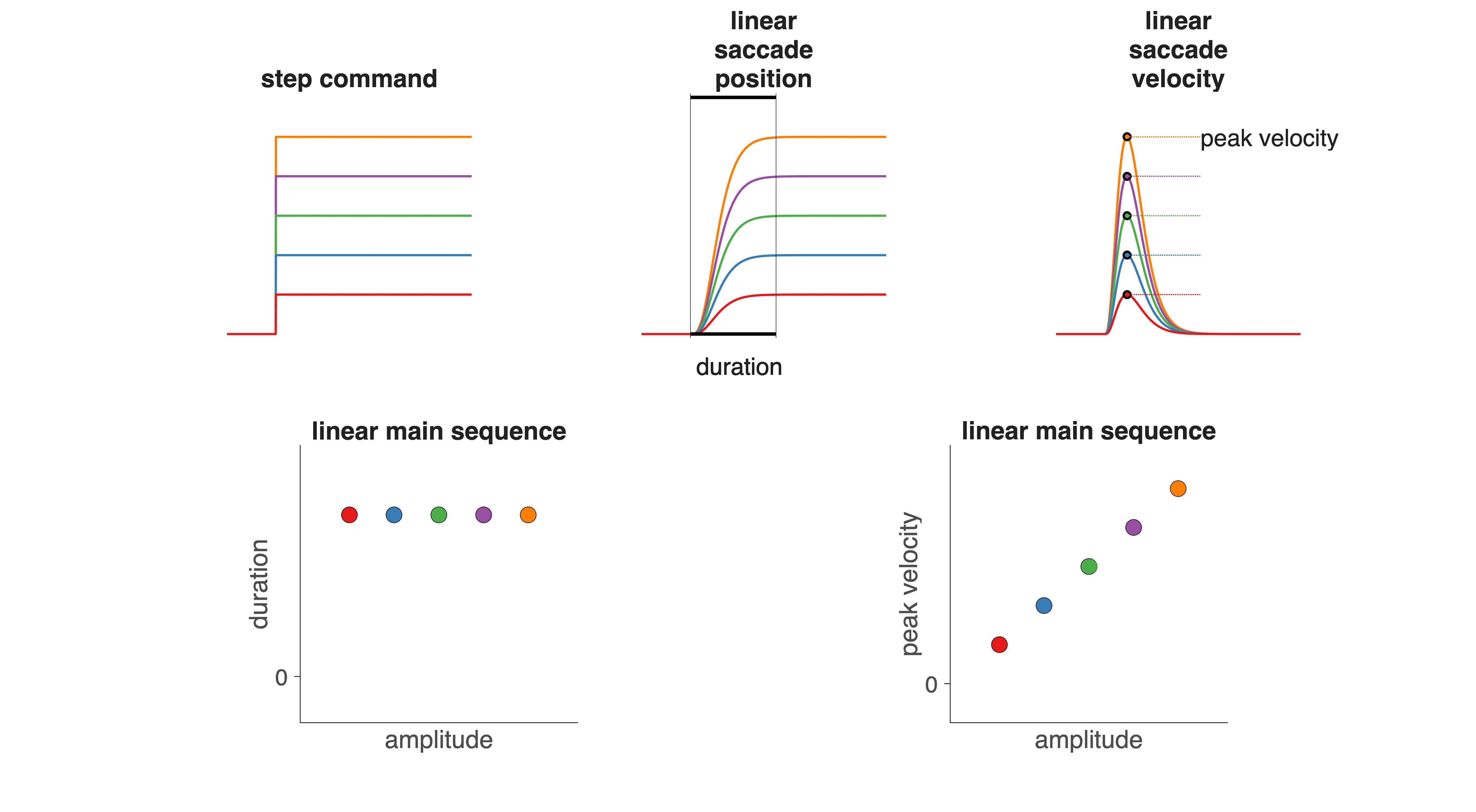
Fig. 42 Hint.#
In a linear (time-invariant) system:
Scaling the input changes only the amplitude of the output.
It does not change the time course (temporal shape) of the response.
Background: Higher-level processes underlying saccade generation
Before the brainstem can generate the eye movement itself, several higher-level processes must occur elsewhere in the brain. These stages are not the focus of this module, but provide useful context.
Target selection The visual scene contains far more potential stimuli than we can look at. The central nervous system therefore must select one location as the goal for the next saccade. This selection is influenced by attention, task demands, and suppression of inappropriate reflexive responses. These mechanisms are largely cortical.
Computation of movement coordinates Once a target is selected, the system must determine the desired saccade vector: the direction and amplitude of the required eye movement. This computation takes into account the retinal error (distance of the target from the fovea), the current eye position in the head, and even the orientation of the head on the neck. This course does not cover these transformations in detail.
Saccade initiation (“triggering”) A decision must then be made to abandon the current fixation and initiate the movement. This involves coordinated suppression of fixation-related activity and activation of movement-related structures. Triggering is a key function of the superior colliculus and associated cortical areas.
Eye-movement control Once the movement begins, an internal feedback mechanism monitors how far the eye has moved and how far it still needs to go. This feedback-based control—implemented by brainstem burst neurons, pause neurons, and the neural integrator—is the focus of the present module.
In simple conditions (e.g., a single target in a dark room), the entire sequence takes roughly 200 ms. In cluttered or ambiguous environments, reaction times increase substantially (see Chapter Reaction Times).
Fig. 43 shows the main cortical and subcortical regions involved in preparing a saccade. Visual information flows from the retina to the lateral geniculate nucleus (LGN), then to primary visual cortex (V1), parietal cortex (PPC), and frontal areas such as the frontal eye fields (FEF) and supplementary eye fields (SEF). These structures contribute to selecting targets, allocating attention, and evaluating the need for an eye movement. Their output ultimately converges onto the Superior Colliculus (SC), a midbrain structure that specifies the desired saccade vector and sends this command to the brainstem, where the actual movement is executed.
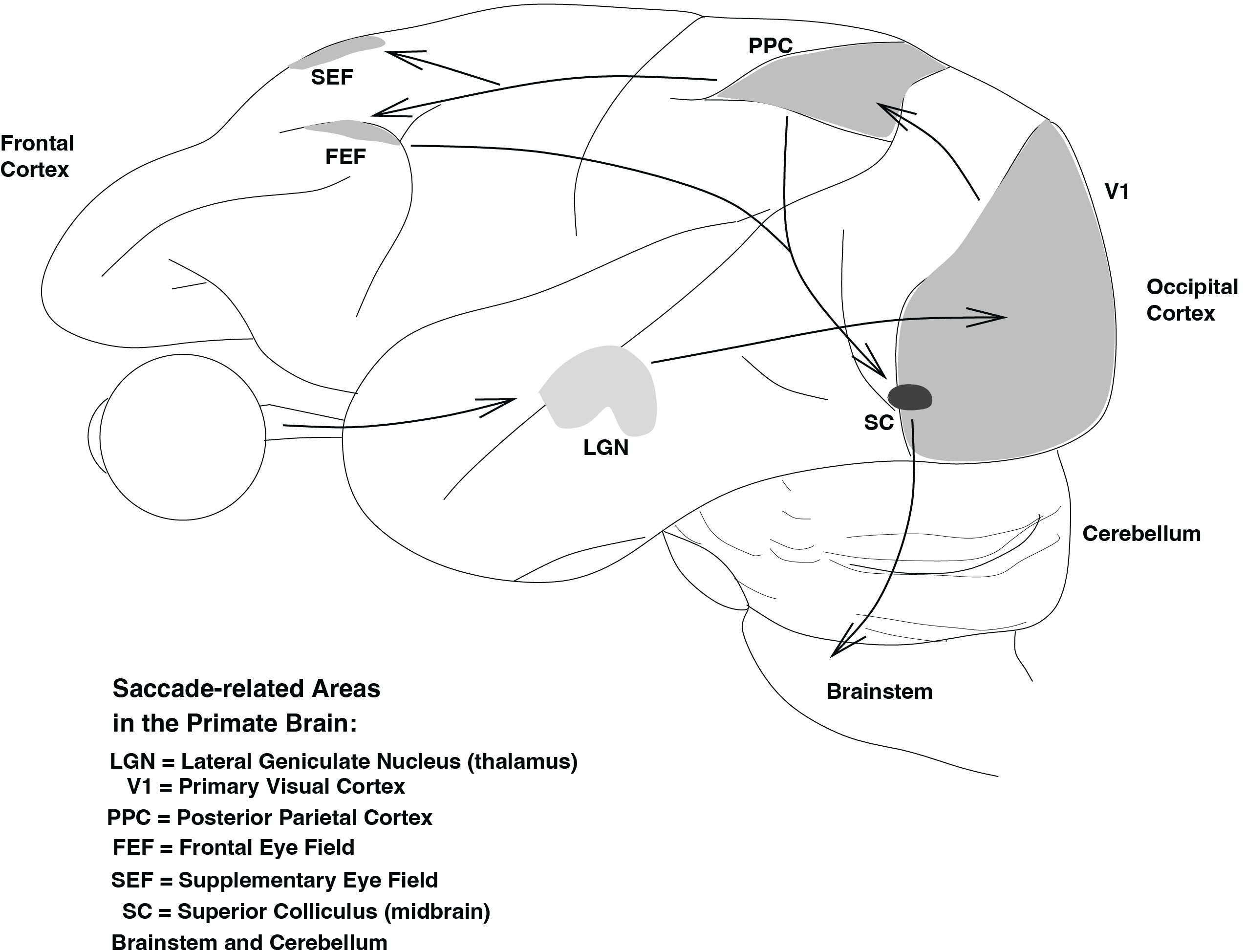
Fig. 43 | Cortex#
Brain regions involved in saccade generation. Only major cortical areas (V1, PPC, FEF, SEF) and subcortical structures (LGN, SC) are shown. The brainstem circuitry responsible for executing the saccade will be discussed later in this module.
Pulse-Step Generator#
From the Main Sequence, we concluded that the oculomotor plant is far too sluggish to produce fast saccades from a simple step input, and therefore must be driven by a pulse–step command: a brief, high-amplitude pulse to rapidly move the eyes, followed by a tonic step to hold them in their new position. We now examine how the nervous system can construct such a command.
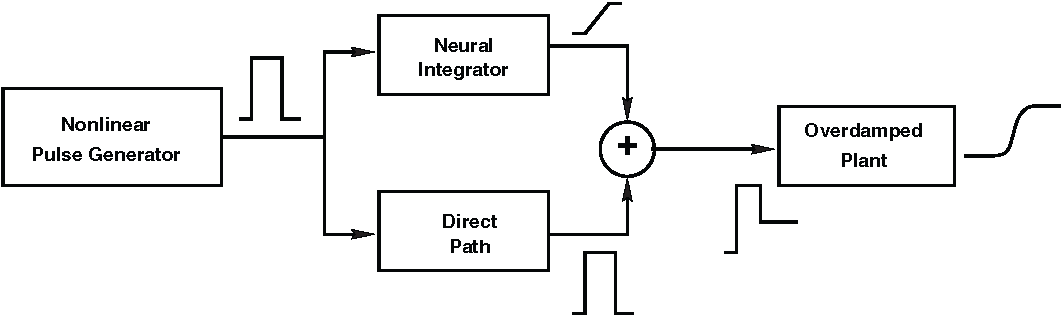
Fig. 44 | Pulse-step generator.#
The schematic pulse–step generator consists of three functional stages:
Pulse generation (burst neurons in the PPRF),
Neural integration of the pulse to form the step (tonic neurons in the NPH), and
Summation of pulse and step at the motoneurons (OMN).
See also the Computer Exercises (Assignment: The Linear Pulse-Step generator) in which this model system is simulated in MATLAB’s simulink.
The system shown in Fig. 44 captures the essential computations needed to transform a desired eye movement into a neural command that can drive the oculomotor plant. It consists of three stages:
Pulse generation
A population of neurons produces a brief, high-frequency burst of activity.
This pulse provides the strong, short-lived force needed to accelerate the eyes rapidly.
Neural integration to create the step
Another population integrates (mathematically sums over time) the pulse signal.
The output is a step: a sustained level of activity proportional to the final eye position.
This integrator compensates for the plant’s elasticity and keeps the eyes at their new position.
Summation of pulse and step
Motoneurons combine the fast pulse and the sustained step into a single command signal.
This composite signal is sent to the extraocular muscles and drives the actual eye movement.
Together, these three operations produce the pulse–step command that is both biologically feasible and mechanically appropriate for moving the eyes quickly and holding them steady.
It is important to emphasize that each of these subsystems — pulse generator, integrator, and summing stage — is linear in this simplified functional model. Linear systems cannot by themselves produce the nonlinear features of the Main Sequence. Thus, while this system explains how a pulse–step command can be constructed, it does not explain the nonlinear dynamics of real saccades. To understand where those nonlinearities arise, we must look upstream, in the circuitry that specifies the desired movement — specifically in the superior colliculus or the burst-neuron nonlinearity. That will be the focus of the next modules (The Saccadic System - The Burst Generator and The Saccadic System - Superior Colliculus) and the practical exercise (Assignment: The Nonlinear Burst Generator).
The Oculomotor Plant#
Linear Model#
In Fig. 44, the final stage of the pulse–step generator feeds into the oculomotor plant: the mechanical system consisting of the eyeball, extraocular muscles, connective tissue, and surrounding orbital mechanics. It is this plant that actually produces the saccadic eye movement in response to the neural command. We can model this plant as a linear system (Fig. 45).
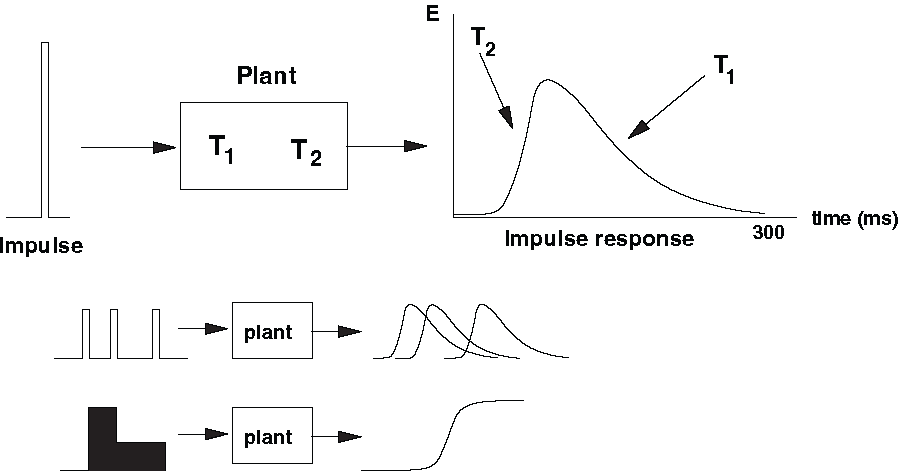
Fig. 45 | Oculomotor plant.#
A linear model of the plant is fully characterized by its impulse response — the miniature eye movement that would result from a single spike entering the motoneurons. Because the plant is (approximately) linear over a wide operating range, the motion produced by any input can be reconstructed by summing scaled and shifted copies of this impulse response (superposition principle).
The plant is highly overdamped#
Mechanically, the plant behaves like a second-order overdamped system. Overdamping means:
it responds slowly and smoothly to a sudden input,
velocity rises gradually and decays gradually,
movements are monotonic (no overshoot).
it does not oscillate or ring.
This is consistent with intuition: pushing the eye with a constant force does not make it “snap” into place — it creeps toward the new position.
Modeled as a second-order linear system#
The plant is well approximated as a second-order linear system. This implies two key properties:
Its input–output behavior can be described by a second-order linear differential equation.
Its impulse response is the sum of two exponential decays, each associated with a time constant.
The two time constants reflect different mechanical aspects of the plant (Fig. 46a):
Long time constant \(T_1 \approx 150\ \text{ms}\)
determined primarily by muscle elasticity (“springiness”)
represents slow settling of the system
Short time constant \(T_2 \approx 12\ \text{ms}\)
related to viscosity (“friction”) of the tissues
represents fast initial response
The impulse response \(h(\tau)\) is given by:
The fast component rises quickly but decays rapidly; the slow component decays much more gradually. Together, they generate a smooth, slow, overdamped movement.
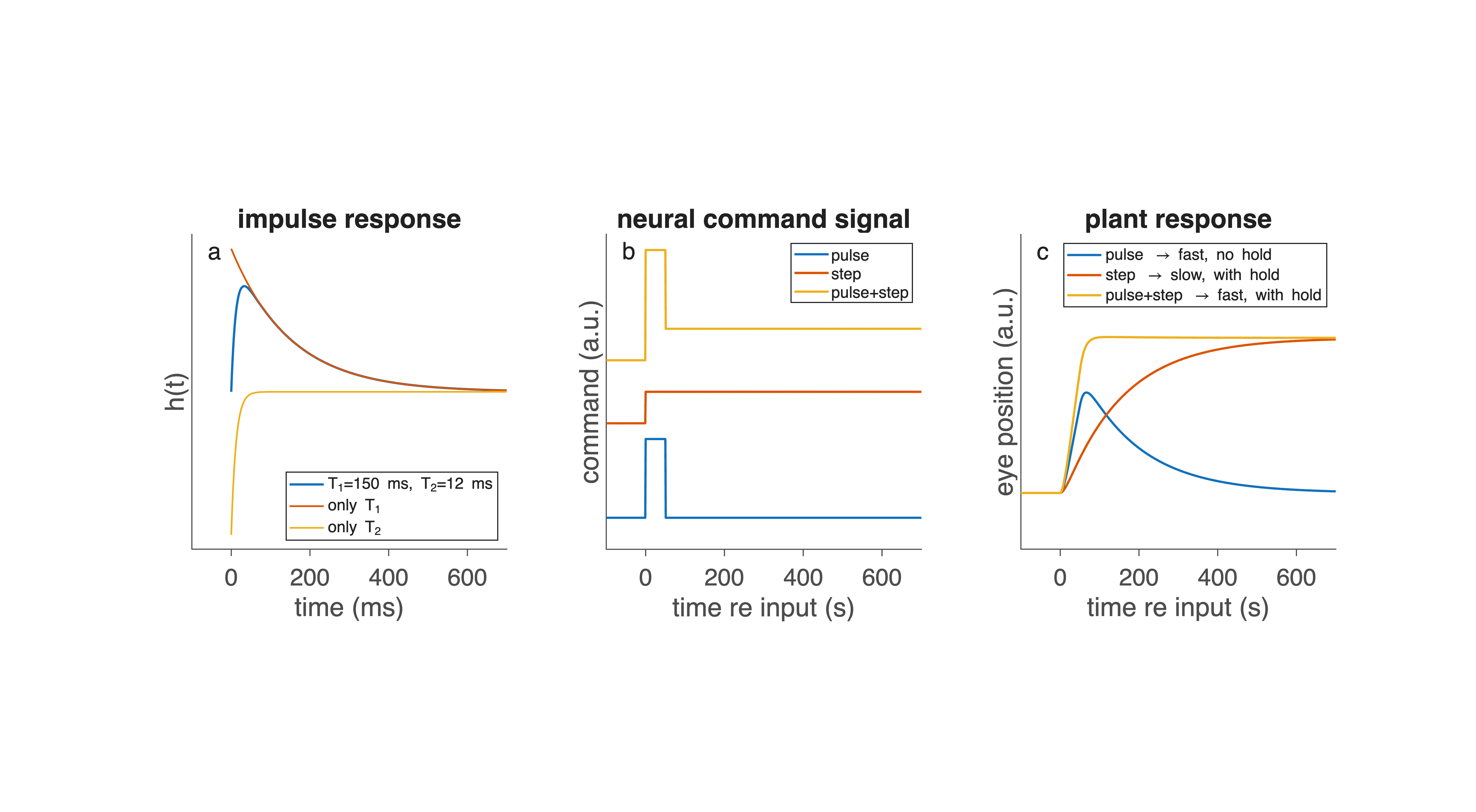
Fig. 46 | Response properties of the oculomotor plant.#
a. Impulse response of the second-order, overdamped oculomotor plant, determined by two time constants: a fast viscous component (T₂) and a slower elastic component (T₁). The effects of selectively altering each time constant are shown for comparison.
b. Three input commands that may drive the plant: a brief pulse, a sustained step, and a combined pulse–step command.
c. Corresponding plant responses. A pulse produces a fast movement without stable fixation; a step produces a slow movement but maintains final position; only the pulse–step command produces both rapid movement and stable fixation — the hallmark of a saccade.
How the plant responds to different inputs#
Because the plant is linear (within a wide range of eye movements), we can predict how it behaves (Fig. 46c) when driven by different types of neural input (Fig. 46b):
Step input → very slow eye movement. A step is a sudden increase in constant muscle activation.
Plant responds with a slow, monotonic exponential movement.
Movement takes hundreds of milliseconds.
Cannot generate fast saccades.
But it does produce fixation once the eye finally arrives.
Pulse input → fast movement but no fixation. A pulse is a brief burst of high activation.
Plant accelerates quickly → rapid eye motion.
But when the pulse ends, activation drops to zero.
Eye drifts back due to muscle elasticity.
Fast but unstable — the eye cannot hold position.
Pulse–step input → fast movement and stable fixation. Combining both inputs gives:
Pulse: provides the rapid acceleration needed for a fast saccade.
Step: holds the eye at its new position after the movement.
This is exactly why the nervous system must generate a pulse–step command: it is the only type of input that both moves the eye quickly and keeps it there.
question – plant as \(2^{nd}\) order linear system
Make a graph of \(h(\tau)\), and determine the location of its maximum.
Solution
For the graph of the impulse response, see Fig. 46a.
If you plot the impulse response in (22) using typical oculomotor values (e.g. \(T_1 = 150\) ms and \(T_2 = 12\) ms), you will find:
\(h(0) = 0\)
the response rises quickly to a single positive peak
and then decays smoothly back to zero
This produces a single-lobed, positive, overdamped impulse response—exactly what we expect from a second-order mechanical system with no oscillations.
A useful way to understand this shape is to look at the two exponential components separately.
At \(\tau = 0\):
the slow exponential \(e^{(−\tau/T_1)}\) starts at +1
the fast exponential \(e^{(−\tau/T_2)}\) also starts at +1, but appears with a minus sign
so they cancel exactly, giving \(h(0)=0\).
Immediately afterward, the fast exponential decays much more rapidly than the slow one. The difference between them quickly becomes positive, causing the impulse response to rise. Once the fast component has nearly vanished, only the slow exponential remains, and the response decays gradually.
You can visualise this behavior by running the MATLAB script
provided in the course,
and inspecting where the peak of \(h(\tau)\) occurs.
For the mathematically inclined
To determine the time τ_max at which h(τ) reaches its maximum, set its derivative to zero:
Setting \(h'(\tau) = 0\) gives:
Taking the natural logarithm:
Thus:
For the typical oculomotor values \(T_1 = 150\) ms and \(T_2 = 12\) ms, this evaluates to:
The Brainstem#
The pulse–step command that drives fast, accurate saccades is generated by a highly organized set of brainstem nuclei. Although many structures contribute to saccade generation, we focus here on the horizontal saccade system, illustrated in Fig. 47. These nuclei together form the final common pathway from the superior colliculus to the extraocular muscles.
The major neuronal populations involved are:
Oculomotor neurons (OMN) in
Abducens nucleus (nVI)
Oculomotor nucleus (nIII)
Tonic neurons in the Nucleus Prepositus Hypoglossi (NPH)
Short-lead excitatory burst neurons (EBN) and inhibitory burst neurons (IBN) in the paramedian pontine reticular formation (PPRF)
Long-lead burst neurons (LLBN) in the PPRF
Omni-pause neurons (OPNs) in the Raphé nucleus
Movement-related neurons in the caudal Superior Colliculus (cSC)
Fixation-related neurons in the rostral Superior Colliculus (rSC)
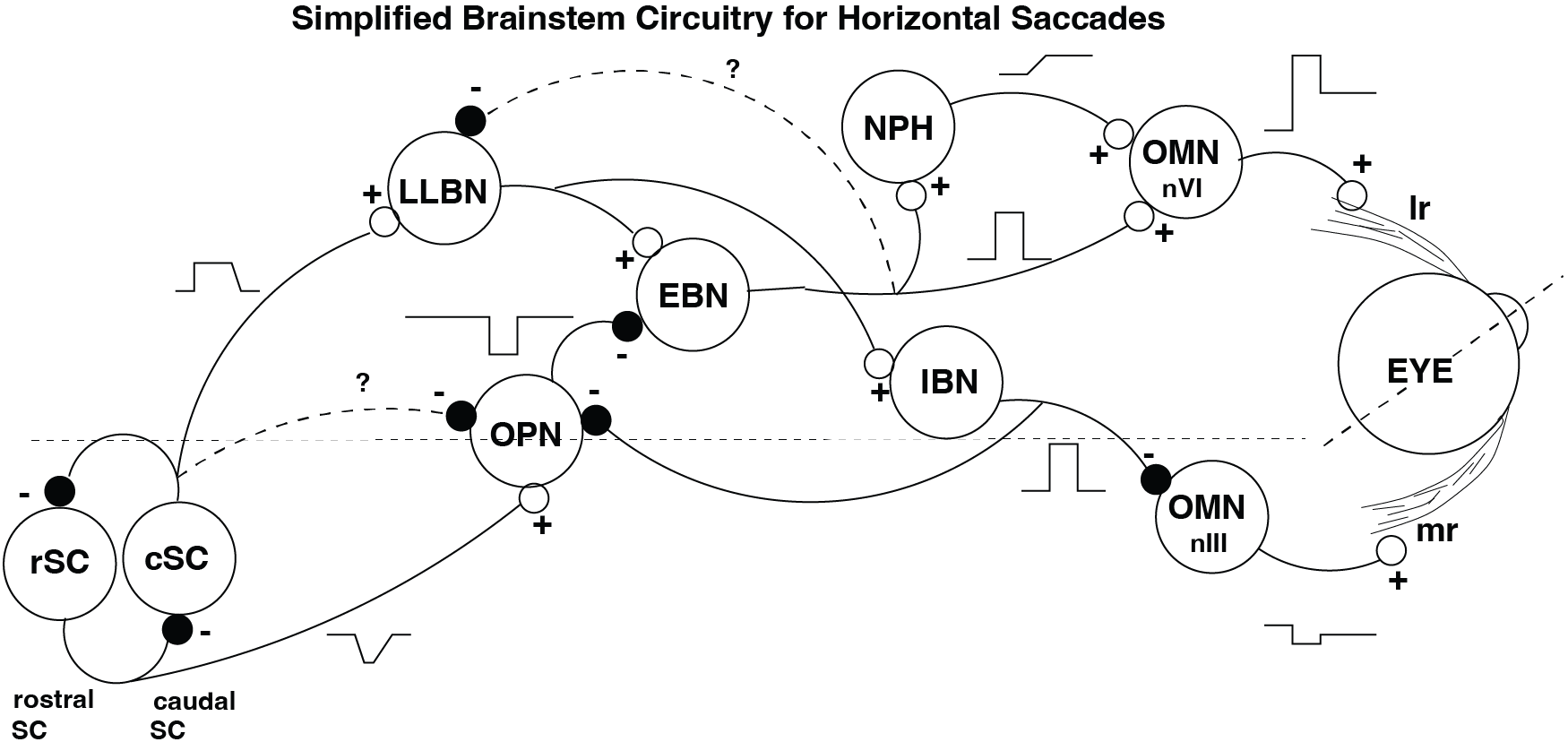
Fig. 47 | Brainstem circuitry for horizontal saccades.#
A simplified scheme of nuclei involved in leftward horizontal saccades (left eye shown). SC = superior colliculus (rostral fixation zone and caudal movement zone), LLBN = long-lead burst neurons, OPN = omni-pause neurons, EBN = excitatory burst neurons, IBN = inhibitory burst neurons, NPH = neural integrator, OMN = oculomotor motoneurons (nVI, nIII). LR = lateral rectus; MR = medial rectus.
Motoneurons: delivering the pulse–step to the muscles#
The oculomotor motoneurons (OMNs) in nVI and nIII send the final command to the eye muscles. Their firing pattern is the pulse–step:
The pulse overcomes the viscous drag at high velocities.
The step holds the eye at its new eccentric position by counteracting elastic restoring forces.
Horizontal saccades require precisely coordinated push–pull activation of different motoneuron pools (Fig. 48) to innervate the eye muscles. For example, horizontal eye movements are generated by either the Lateral Rectus muscle, which pulls each eye outwards, or the Medial Rectus muscle, which pulls each eye inwards. If both eyes are to move rightward, the LR of the right eye, and the MR of the left eye should be activated by a Pulse-Step signal, but at the same time, the MR of the right eye, and LR of the left eye should be silenced:
Agonist muscles (e.g., right LR and left MR for a rightward saccade) → receive the pulse–step command
Antagonist muscles (right MR and left LR for a rightward saccade) → are actively inhibited by IBNs to relax them
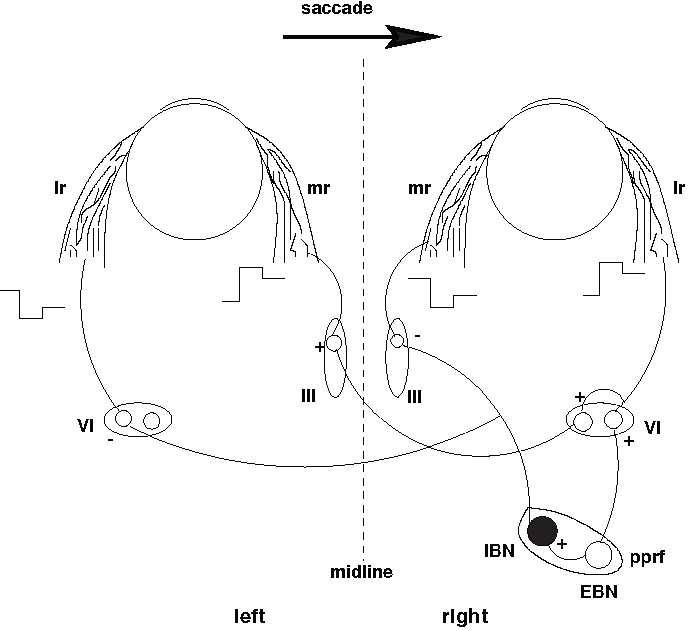
Fig. 48 | Push–pull organization of horizontal motoneuron pools.#
Note the ‘push-pull’ organization of the different motoneuron pools (VI and III).
question – push-pull organization
Explain what happens, when a rightward saccade is made (i.e. the PPRF on the right side sends a Pulse).
Solution
During a rightward saccade, the right PPRF generates a high-frequency pulse.
This activates the agonist muscles that pull the eyes to the right, while simultaneously inhibiting the antagonist muscles that would oppose the movement. This is the classic push–pull organization.
Excitatory pathway (EBNs → OMNs) — drives both eyes to the right
Right EBNs fire a pulse.
They excite:
Right Abducens Motoneurons (nVI) → activates right Lateral Rectus (LR) → pulls right eye outward.
Left Oculomotor Motoneurons (nIII) → activates left Medial Rectus (MR) → pulls left eye inward.
These are the agonist muscles for a rightward saccade.
Inhibitory pathway (IBNs → antagonist OMNs) — relaxes the opposing muscles
The same right EBNs activate right IBNs.
Right IBNs inhibit:
Right Oculomotor Motoneurons (nIII) → relaxes right MR.
Left Abducens Motoneurons (nVI) → relaxes left LR.
These are the antagonist muscles, which must be silenced so they do not oppose the movement.
Summary (push–pull pattern)
Muscle |
Role during rightward saccade |
Effect |
|---|---|---|
Right LR |
Agonist |
Activated (pulls outward) |
Left MR |
Agonist |
Activated (pulls inward) |
Right MR |
Antagonist |
Inhibited (relaxes) |
Left LR |
Antagonist |
Inhibited (relaxes) |
This coordinated activation and inhibition pattern ensures that both eyes move smoothly and rapidly to the right.
question – eye movement disorders
What eye movement abnormalities result after damage of
the Abducens Nucleus (nVI; i.e. the actual cells)?
the Abducens Nerve (i.e. the output of the nucleus to the LR)?
Solution
Damage to the Abducens Nucleus (nVI) and damage to the Abducens Nerve both impair horizontal eye movements, but in different ways.
1. Lesion of the Abducens Nucleus (nVI)
This destroys all neurons in the nucleus:
Abducens motoneurons (which drive the ipsilateral Lateral Rectus)
Abducens internuclear neurons (which project via the MLF to the contralateral Medial Rectus)
Resulting abnormality:
Total loss of horizontal gaze to the lesioned side, affecting both eyes.
The ipsilateral eye cannot abduct (LR paralyzed).
The contralateral eye cannot adduct during conjugate gaze (no MLF internuclear drive to MR).
This is a gaze palsy, not an isolated muscle weakness.
2. Lesion of the Abducens Nerve
This damages only the axon bundle projecting from nVI to the ipsilateral Lateral Rectus.
Resulting abnormality:
Isolated LR paralysis of the affected eye:
The ipsilateral eye cannot abduct.
The contralateral eye behaves normally during horizontal gaze.
Convergence (MR function) is intact.
Eye movement pattern:
Attempt to look toward the side of the lesion → ipsilateral eye stays at midline, contralateral eye abducts normally.
This is a single-eye abduction deficit, not a gaze palsy.
Summary Table
Lesion |
Affected Structures |
Clinical Outcome |
|---|---|---|
Abducens Nucleus (nVI) |
Motoneurons + internuclear neurons |
Horizontal gaze palsy toward the lesion (both eyes affected) |
Abducens Nerve |
LR motor axons only |
Ipsilateral LR paralysis (one eye cannot abduct) |
Burst neurons: generating the pulse#
The Short-Lead Burst Neurons (EBNs and IBNs) in the paramedian pontine reticular formation (PPRF) form the pulse generator.
EBNs excite the agonist motoneurons.
IBNs inhibit the antagonist motoneurons (causing that muscle to relax).
Their firing bursts tightly correlate with eye velocity, and their peak firing rates contribute to the saturation seen in the Main Sequence.
The Long-Lead Burst Neurons (LLBNs) begin firing tens of milliseconds before the saccade and are thought to relay the high-level “go” command from the superior colliculus to the burst generator.
Damage to the PPRF abolishes all horizontal saccades toward the side of the lesion (ipsilateral). This is because the pulse is no longer generated that accelerates the eye to its target position. Even a slow response is missing, as this burst from the EBNs and IBNs is required to produce the step by the neural integrator.
Neural Integrator (NPH): generating the step#
The Nucleus Prepositus Hypoglossi (NPH) contains tonically active neurons whose firing rates encode eye position. This nucleus is widely believed to function as the neural integrator:
It receives the eye-velocity–related burst from EBNs/IBNs
It integrates this signal
Producing the step component sent to motoneurons
A leaky or damaged neural integrator leads to gaze-holding deficits, where the eye drifts back toward center. Note that the pulse is still generated by EBNs/IBNs and so a gaze shift is still produced.
Omni-pause neurons (OPNs): gating the movement#
During fixation, OPNs fire continuously and inhibit EBNs.
Right before a saccade:
OPNs pause, releasing the inhibitory brake on the burst neurons.
When the saccade ends, OPNs start firing again, terminating the burst.
This “pause–burst–pause” interaction ensures that saccades start and stop precisely.
Superior Colliculus (SC): linking intention to action#
Although the superior colliculus is treated in detail in the next module, its two functional zones deserve brief mention here:
The rostral SC (rSC) contains fixation neurons that excite OPNs, preventing saccades.
The caudal SC (cSC) contains movement-related burst neurons whose output encodes the desired saccade vector.
The cSC → LLBN → EBN/IBN pathway forms the bridge from where you want to look to *the brainstem machinery that moves the eye&.
Clinical relevance#
Assuming the oculomotor plant is intact (which is often the case), characteristic eye movement abnormalities (Fig. 49) can be traced back to specific brainstem nuclei.
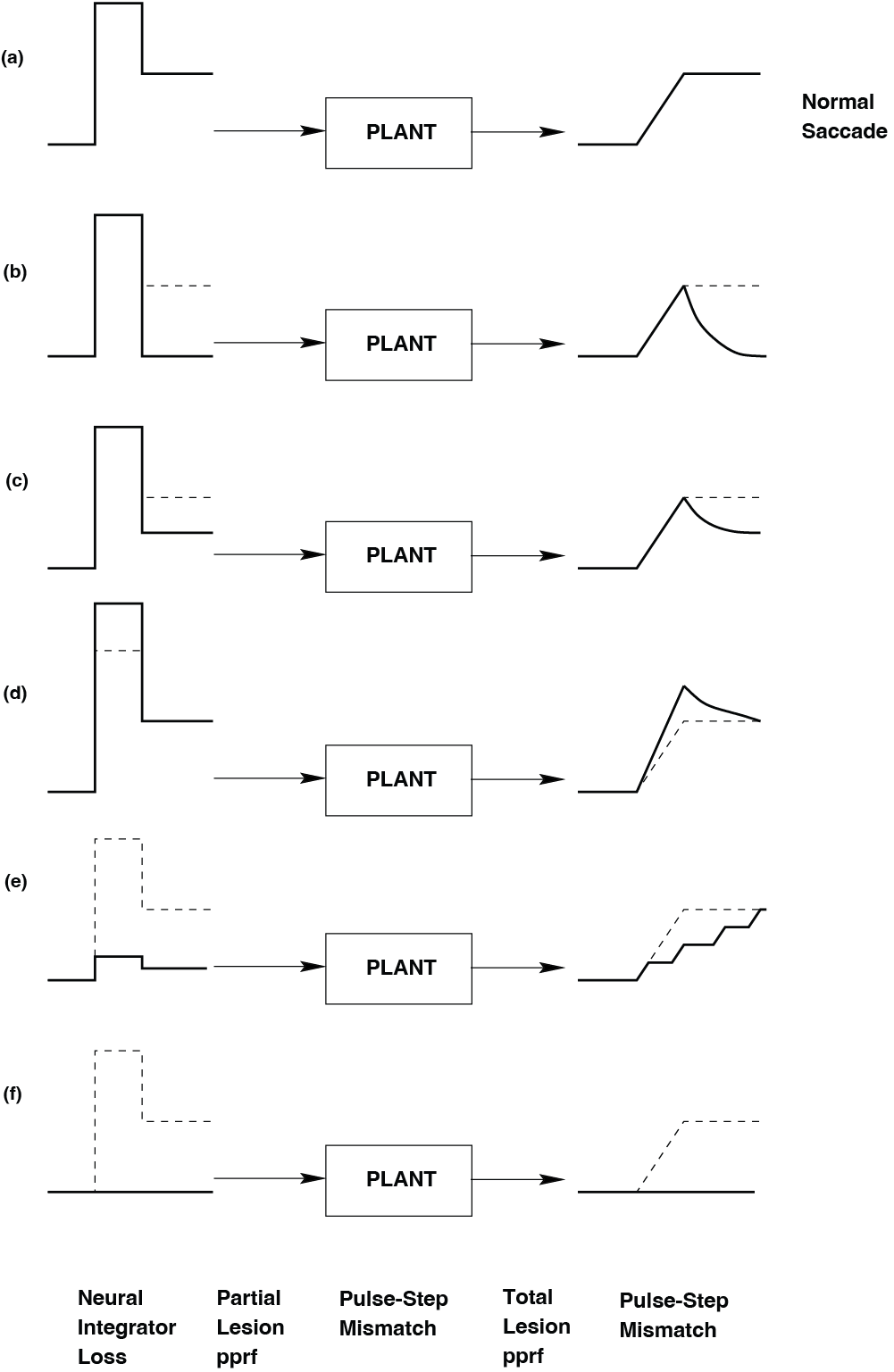
Fig. 49 | Oculomotorpathologies.#
Pathologies in different parts of the pulse–step generator cause distinct abnormalities in saccade behavior.
question – oculomotor pathologies
Assign to each abnormal saccade pattern in Fig. 49 the most likely underlying neural lesion.
Solution
Each panel (a–f) in Fig. 49 shows the eye position trace of a saccade, revealing different abnormalities in the pulse–step generator.
a. Normal saccade
Rapid movement to the target, followed by stable fixation at the new position.
Interpretation: Intact pulse and intact step (no pathology).
b. Loss of the step (no stable fixation) – Neural Integrator Loss
The eye moves rapidly toward the new position, but cannot hold that position.
Instead, it drifts back toward center because the step component is missing.
Interpretation: The neural integrator (e.g. NPH) fails to generate a proper step.
→ Diagnosis: Neural Integrator Loss.
c. Weak step (incomplete fixation) – Pulse–Step Mismatch
The eye initially reaches the approximate target position, but then
drifts partway back, settling at an intermediate, incorrect position.This indicates that the step is present but too small.
Interpretation: The pulse and step are not properly matched in amplitude.
→ Diagnosis: Pulse–Step Mismatch (step too weak).
d. Strong pulse (overshoot) – Pulse–Step Mismatch
The eye overshoots the target position during the saccade, then
comes back toward it (e.g. via a small corrective movement).This suggests a pulse that is too strong relative to the step.
Interpretation: Again a mismatch between pulse and step, but now the pulse is too large (or step too small for the pulse).
→ Diagnosis: Pulse–Step Mismatch (pulse too strong / mismatch).
e. Weak pulse and step (multiple corrective saccades) – Partial Lesion PPRF
The eye moves in the correct direction but with a small, slow saccade,
followed by one or more corrective saccades until it reaches the target.Both pulse and step are present but reduced in size.
Interpretation: The pulse generator is partially damaged; command is weak but not absent.
→ Diagnosis: Partial Lesion PPRF.
f. Absent pulse and step (no saccade) – Total Lesion PPRF
The eye does not move toward the target at all.
No saccade, no pulse, no step.
Interpretation: The saccadic pulse–step generator is completely offline.
→ Diagnosis: Total Lesion PPRF.
Follow-up#
In the computer practical (Assignment: The Linear Pulse-Step generator), we will simulate the pulse-step generator, and try to understand what it does and how it works. In the next module (The Saccadic System - The Burst Generator and The Saccadic System - Superior Colliculus), we will discuss the burst generator, and how saccades are accurately pointing to targets. This module it was about saccades being fast, and how the neural control / command overcomes sluggishness of the plant.
Key terms#
- saccade #
A rapid, ballistic eye movement that shifts the line of sight to a new visual target. Saccades bring objects of interest onto the fovea and can reach peak velocities of 700–1300°/s.
- step response #
The output of a system following a step input. For the oculomotor system, the step response is the saccadic eye movement that brings gaze from the old position to the new target location.
- oculomotor system #
The combined neural and mechanical system that controls eye position and eye movements. It includes the extraocular muscles, the eyeball and orbital tissues (the plant), and the brain structures that generate and shape the motor commands.
- pulse–step command #
The characteristic neural drive that produces saccades a brief, high-frequency pulse that rapidly accelerates the eye, followed by a tonic step of activity that maintains the eye at its new eccentric position.
- reaction time #
The delay between a change in a stimulus (such as a target appearing or jumping) and the onset of the behavioral response. In the context of saccades, it is the time between target onset and saccade onset (also called saccadic latency).
- main sequence #
The stereotyped relationship between saccade amplitude, duration, and peak velocity. In typical Main Sequence data, saccade duration increases approximately linearly with amplitude, and peak velocity increases with amplitude but eventually saturates, revealing nonlinear properties of the saccadic system.
- amplitude #
The size of a movement or change, usually measured in degrees for eye movements. For a saccade, the amplitude is the angular distance the eye travels from its starting position to its final position.
- duration #
The time interval between the onset and the end of a movement or response. For a saccade, the duration is the time between saccade onset and saccade offset.
- peak velocity #
The maximum speed reached during a movement. For a saccade, peak velocity is the highest eye speed reached during the saccade and is a key variable in the Main Sequence.
- plant #
The mechanical part of a control system that is being driven by the controller. In the oculomotor system, the plant consists of the eyeball, extraocular muscles, connective tissues, and orbital mechanics. Because of inertia, viscosity, and elasticity, the plant is naturally too sluggish to produce fast saccades without an appropriately shaped input.
- oculomotor plant #
The oculomotor system’s plant specifically the eyeball, extraocular muscles, and surrounding orbital tissues, considered as a mechanical system driven by neural commands.