The Oculumotor System#
Learning Outcomes#
The saccadic eye movement system is that part of the brain that has the task to redirect the current line of sight as quickly and as accurately as possible to a newly-selected peripheral target. The neural commands to make an eye movement are generated in the Superior Colliculus (this chapter chap:oculomotorsystem) and are relayed to the Brainstem (chapter chap:saccadiccontrol) that consists of an intricate circuit of interconnected nuclei that are responsible for the actual eye movement.
At the end of this section (Part part:saccadicsystem), you will be able to:
Explain how the static model of the superior colliculus generates a saccade
Describe the afferent motor map of the superior colliculus in terms of iso-eccentricity lines, iso-direction lines, complex-logarithmic mapping, retinal and anatomical coordinates, population code, translation-invariant, cell-contribution
Make a drawing of the afferent motor map
Schematically indicate for points in retinal coordinates \(R, \phi\) where they are represented on the motor map
Describe the efferent mapping of the superior colliculus.
Schematically indicate the connection strengths of colliculus cells with the horizontal and vertical oculomotor systems (indicate strength through symbol size)
Explain the shape of movement fields of SC cells for small and large saccades (using the properties of the motor map and the population activity in that map)
Draw the movement fields for amplitude and direction (indicating appropriate variables)
General Background#
This course will cover the saccadic eye movement system, a relatively simple system tasked with redirecting the line of sight to a peripheral target as quickly and accurately as possible. We perform these eye movements about 3 times per second when scanning our visual environment or reading. To provide context, we will review the main brain areas involved in saccadic eye movement generation in primates, including monkeys and humans.
While the cortex will not be specifically addressed in this course, Fig. Brain regions involved in the generation of saccadic eye movements in monkeys. Only the primary cortical areas (V1, PPC, FEF, and SEF) and a few subcortical structures (LGN and SC) are depicted. The intricate network of saccade-related nuclei in the brainstem will be discussed later in this course. The arrows indicate the flow of visual information, starting from the retina to the LGN (a relay station) then to V1, followed by the parietal cortex (PPC) and frontal areas (FEF and SEF). Both the parietal and frontal cortex project to the SC, either directly or through the Substantia Nigra, which then connects to the brainstem. illustrates the cortical areas involved in saccades. The output from several cortical areas is transmitted to the Superior Colliculus, a small (approx. 3x3x2 mm) nucleus in the midbrain that generates the command for the saccadic eye movement. The Superior Colliculus also determines the coordinates of the eye movement, such as how far to the right/left and up/down the eye needs to move to look at the target. These command signals are then relayed to the Brainstem, where an interconnected network of nuclei is responsible for executing the eye movement. This final stage will be the focus of this brief course.
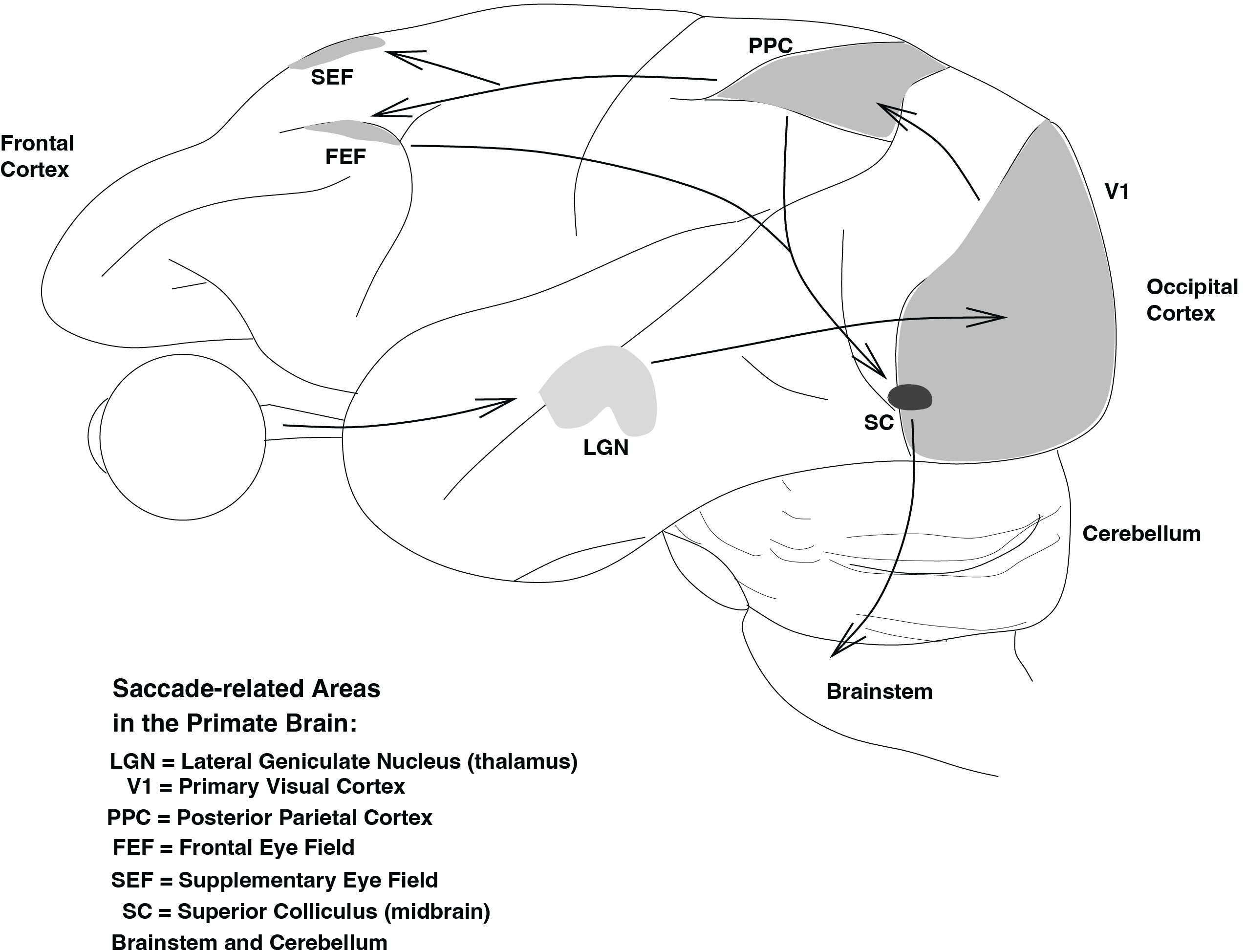
Fig. 36 Brain regions involved in the generation of saccadic eye movements in monkeys. Only the primary cortical areas (V1, PPC, FEF, and SEF) and a few subcortical structures (LGN and SC) are depicted. The intricate network of saccade-related nuclei in the brainstem will be discussed later in this course. The arrows indicate the flow of visual information, starting from the retina to the LGN (a relay station) then to V1, followed by the parietal cortex (PPC) and frontal areas (FEF and SEF). Both the parietal and frontal cortex project to the SC, either directly or through the Substantia Nigra, which then connects to the brainstem.#
Saccades#
Kinematics#
Saccades are rapid and precise eye movements that redirect the line of sight to a new target location outside the current fixation area. These eye movements are an essential aspect of visual processing, allowing us to efficiently scan our environment, read, and attend to moving objects. These fast and precise eye movements reach speeds of up to 700 degrees per second in humans and 1300 degrees per second in rhesus monkeys. From a Systems Theory perspective, the input to the saccadic system can be viewed as a step function (a sudden displacement of the target to a new location), making the saccadic eye movement the step response of the system.
When prompted by an unexpected target, saccades display stereotypical kinematics, or temporal properties, known as the Main Sequence of saccadic eye movements. The Main Sequence of saccades refers to the characteristic relationship between saccade amplitude, duration, and peak eye velocity, as illustrated in Fig. Saccadic Eye Movement Kinematics with Varying Amplitudes. The duration of saccades increases linearly with amplitude, while peak velocity has a saturation relationship. The velocity profile shape varies with amplitude.. The Main Sequence are two graphs: one that plots the relationship between the amplitude of the saccade (the distance the eye moves) and its duration (the time it takes to complete the movement), and the other includes the peak eye velocity (which represents the fastest point during the movement). This relationship is consistent across individuals and species, with the amplitude and duration increasing and decreasing proportionally, respectively, as the saccade becomes larger.
Fig. 37 Saccadic Eye Movement Kinematics with Varying Amplitudes. The duration of saccades increases linearly with amplitude, while peak velocity has a saturation relationship. The velocity profile shape varies with amplitude.#
The Main Sequence is significant because it provides insight into the underlying neural mechanisms responsible for the control of eye movements. The consistent relationship between amplitude, duration, and peak velocity suggests that these eye movements are generated by a highly integrated and optimized system. In addition, the Main Sequence provides a useful tool for researchers and clinicians, allowing them to quantify and compare the properties of saccadic eye movements across different conditions and populations.
From the kinematics and the main sequence, we can already see some interesting features of oculomotor control.
A step-like motoneuron command is insufficient.
Fig. 38 Saccadic Eye Movement Kinematics with Varying Amplitudes. The duration of saccades increases linearly with amplitude, while peak velocity has a saturation relationship. The velocity profile shape varies with amplitude.#
A Pulse-Step motoneuron innervation is needed.
Fig. 39 A pulse-step response of the oculomotor plant (dotted line) may account for the kinematics of saccades.#
The saccadic system is nonlinear!
Fig. 40 Property of a linear system.#
As indicated in Fig. Property of a linear system., linearity of a system requires that whenever the input amplitude of a signal (e.g. the size of the target jump) is multiplied by a certain factor, say a, the system also scales its output by the same factor (see also eqn. eqn:superpositionprinciple):
Therefore, the main sequence belies a nonlinearity in the saccadic system because a linear relationship would mean that peak velocity would increase proportionally with amplitude, and that duration would remain fixed, but this is not the case. Instead, the saturation relationship suggests that some nonlinear mechanism is limiting the ability of the eye to move faster for larger amplitudes, indicating that the saccadic system has some intrinsic nonlinearities. In other words, if the input to the saccadic system is considered to be a Step-displacement of the target on the retina, one should conclude that on the basis of the main-sequence properties of saccades (Fig. Saccadic Eye Movement Kinematics with Varying Amplitudes. The duration of saccades increases linearly with amplitude, while peak velocity has a saturation relationship. The velocity profile shape varies with amplitude.) the saccadic system as a whole is a nonlinear system.
\begin{exercise}[Linear Main Sequence] Why is this conclusion drawn? Draw the expected Main Sequence of a (i.e. any!) linear system.
\end{exercise}
Processes underlying saccade generation#
Target Selection: The central nervous system (CNS) must select the target for the saccade from an almost infinite number of potential visual and auditory targets. Reflexes need to be avoided. Instead, attentional (higher, cortical) mechanisms play a crucial role in this selection process.
Computation of Eye Movement Coordinates: The saccadic system must calculate the direction and amplitude of the ensuing eye movement by considering not only the retinal error vector of the target (i.e. the angular distance of the target from the fovea), but also the starting position of the eyes in the head and the orientation of the head on the neck. This aspect is not covered in the course.
Saccade Initiation: The actual decision to initiate the saccade, also known as the “trigger,” must be made, and the current fixation must be “disengaged.”
Eye Movement Control: An internal feedback loop is used by the saccadic system to continuously compare the actual eye movement to the desired eye movement. This stage, which is the main topic of this section, will be discussed in further detail.
In a simple visual environment, such as a single target in a dark room, these stages typically take about 200 ms to initiate a saccade. However, in more complex visual scenes, the reaction time (or latency) rapidly increases (see also Chapter chap:reactiontimes).
The Brainstem#
A number of different cell groups in the brainstem and midbrain have been extensively studied and much is known about their role in saccades. Here, we will limit the discussion to horizontal saccades (see Fig. Brainstem. A highly simplified scheme of the brainstem regions involved in the generation of horizontal (leftward) saccadic eye movements, shown for the left eye only. SC = superior colliculus (rostral zone and caudal part), LLBN = long-lead burst neurons, OPN = omni-pause neurons, EBN = shortlead excitatory burst neurons, NPH = tonic neurons, OMN = oculomotor neurons (nVI = abducens nucleus, nIII = oculomotor nucleus), IBN = inhibitory burst neurons. LR = lateral rectus muscle, MR = medial rectus muscle. Not all inhibitory interneurons are shown, for reasons of clarity.), for which the relevant cell groups are (read this, learn it by heart, and you can have intelligent discussions with any neurologist):
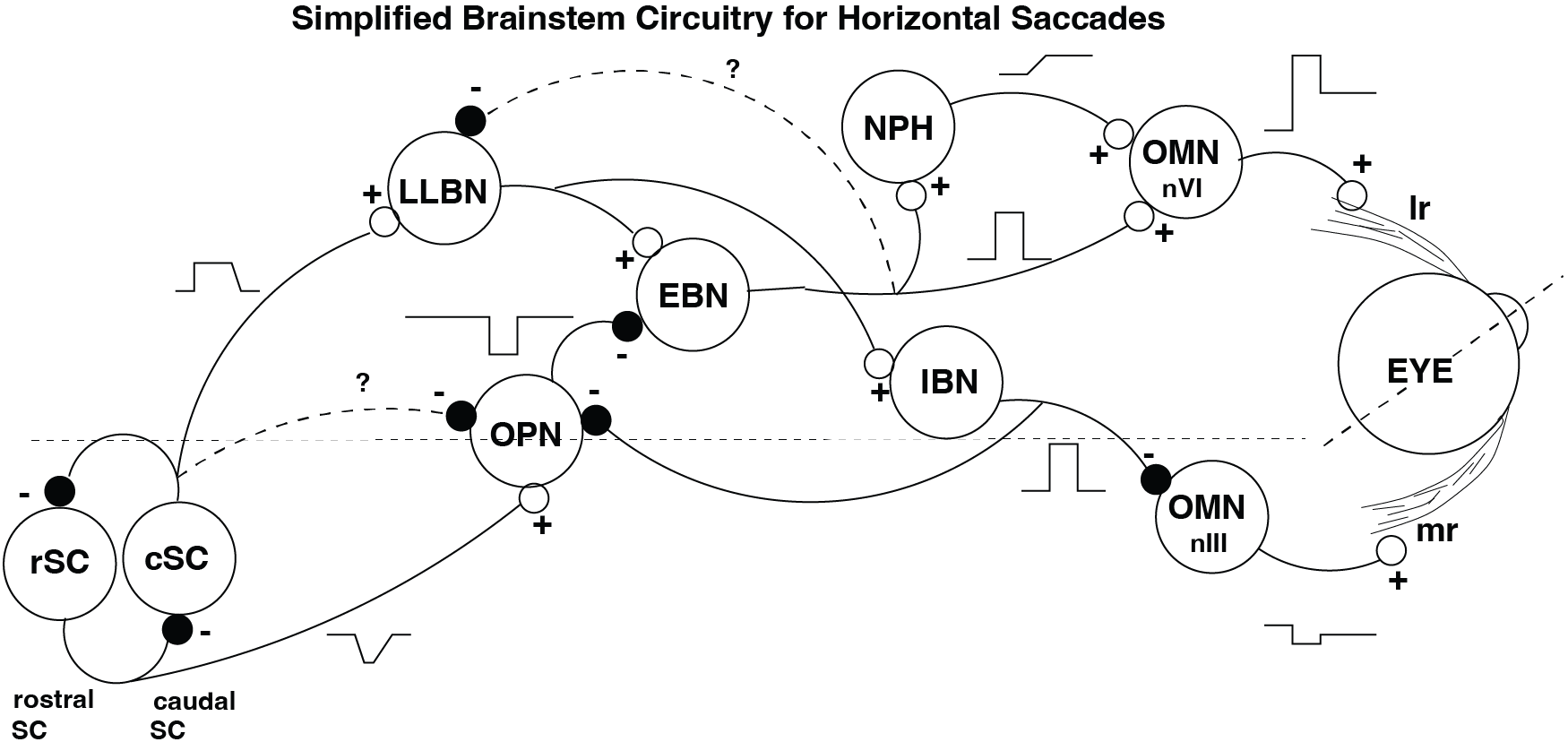
Fig. 41 Brainstem. A highly simplified scheme of the brainstem regions involved in the generation of horizontal (leftward) saccadic eye movements, shown for the left eye only. SC = superior colliculus (rostral zone and caudal part), LLBN = long-lead burst neurons, OPN = omni-pause neurons, EBN = shortlead excitatory burst neurons, NPH = tonic neurons, OMN = oculomotor neurons (nVI = abducens nucleus, nIII = oculomotor nucleus), IBN = inhibitory burst neurons. LR = lateral rectus muscle, MR = medial rectus muscle. Not all inhibitory interneurons are shown, for reasons of clarity.#
Oculomotor neurons in the Abducens Nucleus (n.VI) and the third nucleus (n.III) (OMN)
Tonic neurons (i.e. neurons firing at a d.c.-rate) in the Nucleus Prepositus Hypoglossi (NPH)
Short-Lead Excitatory (EBN) and Inhibitory (IBN) Burst neurons in the paramedian pontine reticular formation (PPRF).
Long-Lead Burst Neurons in the PPRF (LLBN)
Omni-pause neurons in the Raphe Nucleus (OPN)
Saccade-related burst neurons in the caudal Superior Colliculus (cSC)
Fixation-related neurons in the rostral (=‘front’) portion of the SC (rSC).
Fig. 42 Pulse-step generator. The pulse-step generator of the saccadic system involves three stages (neural populations): pulse generation (PPRF), neural integration (NPH), and summation (OMN). See also the Computer exercises.#
The Oculomotoneurons (OMN) control a heavily-damped ‘plant’ (short-cut name for the ocular globe, the six extraocular muscles and surrounding tissues; see also below) with a so-called Pulse-Step command signal on the agonist muscle (see above), and often a complete cessation of activity on the antagonist (Note: a muscle’s action is called agonistic when it pulls into the direction of net motion; it’s called antagonistic when it pulls in the opposite direction.) The Pulse component of this signal is needed to overcome the relatively strong viscous (‘friction’) forces that are especially prominent at the high saccade velocities (note that friction increases roughly quadratically with velocity; Figs. A spring-mass system exhibiting undamped (top) and damped (bottom) oscillations.). The Step component of the signal keeps the eye in its new peripheral orientation after the saccade has ended. It thus precisely compensates for the static elastic restoring forces of the eye muscles (which behave like mechanical ‘springs’). Note, that the different neuronal pools that have to innervate the eye muscles, should be precisely coordinated. For example, horizontal eye movements are generated by either the Lateral Rectus muscle, which pulls each eye outwards, or the Medial Rectus muscle, which pulls each eye inwards. If both eyes are to move rightward, the LR of the right eye, and the MR of the left eye should be activated by a Pulse-Step signal, but at the same time, the MR of the right eye, and LR of the left eye should be silenced! Below, in Fig. Oculomotoneuron control of both eyes for horizontal saccades. Note the ‘push-pull’ organization of the different motoneuron pools (VI and III). Explain what happens, when a rightward saccade is made (i.e. the PPRF on the right side sends a Pulse)., a more complete scheme is shown that does exactly that:
Fig. 43 Oculomotoneuron control of both eyes for horizontal saccades. Note the ‘push-pull’ organization of the different motoneuron pools (VI and III). Explain what happens, when a rightward saccade is made (i.e. the PPRF on the right side sends a Pulse).#
\begin{exercise}[Eye movement disorders] What eye movement abnormalities result after damage of
[label=(\alph*)]
the Abducens Nucleus (nVI; i.e. the actual cells)?
the Abducens Nerve (i.e. the output of the nucleus to the LR)?
\end{exercise}
The Long-Lead and Short-Lead Burst Neurons are a crucial part of the central saccade generator in the brainstem. Lesions (=‘damage’) in the paramedian pontine reticular formation, where these neurons are found, lead to a complete abolishment of all horizontal saccades toward the side ipsilateral to (i.e. to the same side of) the lesion. It is generally held that these neurons are crucial for the generation of both the pulse and the step components of the motoneuron command for both eyes. The antagonist muscle relaxes due to the action of inhibitory burst neurons (IBNs; Fig. Oculomotoneuron control of both eyes for horizontal saccades. Note the ‘push-pull’ organization of the different motoneuron pools (VI and III). Explain what happens, when a rightward saccade is made (i.e. the PPRF on the right side sends a Pulse).). Quantitative analysis of the activity of these burst neurons (especially in the EBNs and IBNs) shows a strong relation with eye velocity, and it is thought that these firing patterns underly the nonlinearity in saccade kinematics (see above). This relation is less tight in the LLBNs.
The tonically active neurons in the NPH are supposed to embody the neural integrator that generates an eye-position related signal (the ‘step’) from the velocity burst of the EBNs/IBNs.
Fig. 44 Oculomotor plant. Linear model of the plant is fully characterized by its impulse response. Conceptually, the impulse response is the (miniature) eye movement that would result in response to a single spike at its input. Due to the **superposition principle#
The ‘plant’ (Fig. {ref}`fig:oculomotorplant**) is highly over-damped, but linear for a considerable range of movements. It is usually modeled as a second-order linear system. This means two things:
that its input-output relation can be described by a second-order linear differential equation, and
that its impulse response can be characterized by two time constants.
The long time constant (\(T_1\) about 150 ms) is mainly determined by the elastic properties of the muscles (the spring constant), the short time constant (\(T_2\) about 12 ms) is related to the plant’s viscosity (‘friction’). The Impulse response of the plant, \(h(\tau)\), is given by the following equation: ```{math` :label: eqn:plant2ndordereqn h(\tau) = \frac{1}{T_1-T_2}[\exp(\frac{-\tau}{T_1})-\exp(\frac{-\tau}{T_2})]
\begin{exercise}[Plant as 2\textsuperscript{nd} order linear system]
Make a graph of $h(\tau)$, and determine the location of its maximum.
\end{exercise}
By assuming that in certain neurological diseases the oculomotor plant itself remains unaffected, many typical oculomotor problems associated with horizontal saccades can be diagnosed.
The **OmniPauseNeurons** (OPN) are crucially involved in the **initiation** of the saccade. Their firing follows a reverse pattern as that of burst neurons: they are active during fixation, and silent during the saccade. It is therefore thought that EBNs and OPNs form mutually inhibitory connections.
As has been extensively documented by a number of studies, cells in the caudal zone of the **Superior Colliculus** are arranged in a so-called **Motor Map**. Each saccade vector appears to be associated with a specific group of
SC cells. Such a localized group of cells sends a burst of spikes to the brainstem saccade generator. Burst onset is tightly coupled to saccade onset, but only loosely related to saccade kinematics. In fact, the burst properties of different SC sites appear to be very similar across the entire motor map. In models of the SC it is therefore assumed that large and small saccades are encoded by the synaptic connectivity of the different groups of SC cells, rather than by their specific firing rates. Cells in the rostral pole of the SC having firing properties that resemble those of the OPNs, to which they make direct connections. It is therefore assumed that these rostral cells play a role in fixation by exciting the OPNs.
```{figure} images/pathologies.pdf
:name: fig:pathologies
Oculomotorpathologies. Pathologies in different parts of the pulse-step generator cause specific abnormalities in saccade behavior.
\begin{exercise}[Oculomotor pathologies] Assign, in Fig. fig:pathologies, the correct neural pathology to its associated saccade abnormality. \end{exercise}
Fig. 45 The superior colliculus motor map. A local population of cells in the caudal SC motor map (circular, grey-shaded area) sends a (roughly fixed) burst of spikes (top-left figure) to the brainstem. A given population encodes a fixed eye displacement vector, irrespective of initial eye position (right figure). Thus, for all saccades shown, the same population of cells will be equally recruited. Different saccade displacements are encoded by different populations in the map. r=rostral fixation zone.#
A Quantitative Model of the Saccadic System.#
Fig. 46 Saccade control model. This includes the Superior Colliculus (SC), the Long-Lead Burst neurons (LLB) and the burst generator in the brainstem. The saccade Gate (OPN firing rate) is controlled by the rostral part of the Superior Colliculus: its fixation-related activity (and thus EBN inhibition) prevents saccades. The saccade trigger (which inhibitis the OPNs, thus releasing the EBNs) is given by the caudal SC cells in the motor map.#
In this part, a conceptual saccade-control model is discussed. This theoretical model is based on a vast amount of experimental data (obtained from trained monkeys and cats). Due to the relative simplicity of this model, and to its close correspondence to realistic midbrain-brainstem neurophysiology, it is a nice model to illustrate some of the basic concepts that have been dominating the oculomotor field during the last two decades. The model is also a fine example of the power of computational modeling with Neural Networks and System’s Theory and of their potential to probe and understand neurobiological systems and pathologies. The model provides a detailed explanation of the two final steps in the saccade generation process (see above): the initiation and the execution of a saccadic eye movement. Note, that the model is not intended to mimic cognitive behavior, or target selection mechanisms (like the Countermanding Paradigm or Antisaccade Paradigm). Everything considered ‘upstream’ from the Superior Colliculus (Basal Ganglia and Cortex) is beyond the scope of the model. However, the model does provide insight into several properties of the saccadic system (see above), and of the connectivities and functional role of the midbrain Superior Colliculus and of several crucial brainstem nuclei. It is also very useful for the diagnosis of certain neurological pathologies that appear to have their effects on saccade generation mechanisms (such as brainstem palsies, Parkison’s disease, Huntington’s disease, etc.). Several of these aspects may be studied by means of the computer simulation programs. The model is based on a number of assumptions and experimental findings:
Saccades are controlled by an internal feedback loop. In the model, the feedback loop carries the velocity output from the Inhibitory Burst Neurons. Below, we will discuss the potential benefits of feedback into somewhat more detail.
The Long-lead Burst cells compare the saccadic command from the Superior Colliculus, with the neural (internal feedback) estimate of the actual eye movement. The difference between these signals is timeintegrated by the LLBNs. Their output is a neural estimate of the current motor error, me. The motor error is the distance the eye still is to go to reach the target. This signal drives the two kinds of short-lead burst cells (EBNs and IBNs).
The Burst Neurons have a nonlinear (saturating) input-output characteristic (Fig. 15). In the saccade model, the relation between current motor error and the eye-velocity (pulse) command is given by:
\[\]
:label: eqn:bniochar v(t) = V_0 \cdot [1 - e^{\frac{me(t)}{e_0}}]
where $V_0$ is the asymptote, and $e_0$ is the angular constant of the nonlinearity.
```{figure} images/interruptedsaccade.pdf
:name: fig:interruptedsaccade
Evidence for internal feedback in the saccadic system. Brief stimulation of the OPNs causes the eye to stop in midflight. However, in the absence of visual feedback, the resuming eye movement reaches the extinguished target position.
Fig. 47 The LLBNs integrate the ongoing difference between the SC burst and the feedback velocity signal from the EBNs. Note, that according to the saccade model, the SC firing rate encodes desired eye velocity, whereas the number of spikes in the SC burst encodes the desired eye displacement, \(\Delta E\). The output of the LLBNs is instantaneous eye motor error, \(me(t)\).#
Fig. 48 The EBNs and IBNs embody the saccadic pulse generator. It transforms eye motor error into an eye velocity command: the larger eye motor error, the higher the eye velocity. As motor error falls to zero (due to the internal feedback loop), eye velocity goes to zero too. Note that the transformation is a saturating nonlinearity, that accounts for the Main Sequence of Fig. {ref}`fig:mainsequence#
Feedback#
Feedback is an important feature of many biological systems. Since it occurs so commonly, there must be a good reason for such an organization. Let’s look at a very simple example, which illustrates the main basic properties. The negative feedback (why?) system depicted in Fig. 16 contains two linear gain elements (both real and positive). The input of the system is first compared with an amplified version of the output, before it is being transmitted through the forward gain.
```{figure` images/simplefeedback.pdf :name: fig:simplefeedback TA simple feedback system, with forward gain, G, and feedback gain, H.
\begin{exercise}[Gain of simple feedback system]
Verify that the relation between input and output (i.e. the overall gain of the system) is now given by:
```{math}
:label: eqn:gainfeedback
\frac{Y(t)}{X(t)} = \frac{G}{1+G\cdot H}
\end{exercise}
\begin{remark} If \(G \cdot H >> 1\) , the transfer of the system is given by \(\frac{1}{H}\), which is independent of the gain \(G\). In this way, the feedback system as a whole can be made very reliable, despite the fact that part of its elements may be prone to damage, or noise. If \(H = 1\) (direct feedback), and \(G\) is sufficiently large, the overall gain of the system can get arbitrarily close to 1, again independent of the actual value of \(G\). \end{remark}
\begin{exercise}[Positive feedback] What happens in the case of positive feedback? \end{exercise}